

Partie 1
Réception : les trois oreilles
B. Conversion en signal électrique — l'oreille interne

[Fig. 2.9] Schéma de la situation de l'oreille interne par rapport aux trois oreilles
(Cliquez pour voir l'original)
1) Structure générale
L’oreille interne est composée de deux parties principales appelées respectivement labyrinthe osseux et labyrinthe membraneux.
Le labyrinthe osseux est une cavité creusée directement dans l’os temporal. Sa forme singulière est composée des canaux vestibulaires, sortes d’arcs impliqués dans la localisation dans l’espace, et de la cochlée, tube enroulé sur lui-même qui analyse et transmet les sons au cerveau. Ce labyrinthe osseux est rempli d’un fluide de couleur bleue nommé la périlymphe.
Le labyrinthe membraneux est une enveloppe membraneuse, et donc souple, en suspension dans la périlymphe du labyrinthe osseux. Sa forme est sensiblement la même que celle du labyrinthe osseux. Il est rempli d’un autre fluide bleu, plus visqueux que la périlymphe, appelé endolymphe.
2) La cochlée
Appelée également limaçon, elle s’apparente à une spirale enroulée sur elle-même. Elle est séparée en trois cavités différentes : la rampe vestibulaire, par où l’onde monte dans la cochlée, la rampe tympanique, par où l’onde redescend de la cochlée, et enfin le canal cochléaire, où l’onde est traitée et transformée en signal électrique. Le sommet de la cochlée est appelé l’hélicotrème, ou apex. À ce niveau, les rampes tympaniques et cochléaires se rejoignent (voir figure 2.10).
|
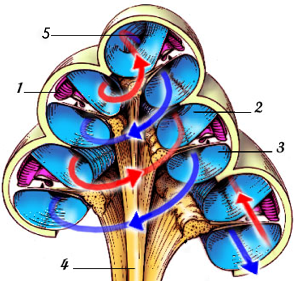 [Fig. 2.10] Schéma d'une coupe de la cochlée représentant le parcours d'une onde à l'intérieur (Cliquez pour voir l'original) |
C’est également au niveau de la cochlée que le son est transformé en signal bioélectrique, et plus précisément dans le canal cochléaire, où se situe l’organe spiral ou organe de Corti, du nom de celui qui le décrit en premier, en 1851 : Alfonso Corti.
3) Conversion de l'onde — Organe de Corti
L’organe de Corti est un tout petit organe composé principalement de cellules sensorielles situé dans le canal cochléaire (voir figure 2.11). Il repose sur la membrane basilaire, en contact avec la rampe tympanique. Son fonctionnement repose sur la friction de cellules ciliées contre la membrane tectoriale : l’onde, en passant dans la rampe vestibulaire, engendre une pression contre la membrane basilaire, laquelle bouge alors de bas en haut, produisant l’inclinaison des cellules ciliées.

[Fig. 2.11]Schéma de l'organe de Corti sur une coupe de la cochlée
(Cliquez pour voir l'original)
4) Les cellules ciliées : de l'onde mécanique à l'impulsion électrique
Les cellules ciliées sont des cellules sensorielles situées sur la couche supérieure de l’organe de Corti. À leur sommet se trouvent des organites particuliers en forme de cils qui sont sensibles aux mouvements de leur environnement. Ces cils baignent dans l’endolymphe, un fluide très concentré en ions potassium K+, et certains sont en contact avec la membrane tectoriale (voir figure 2.12).
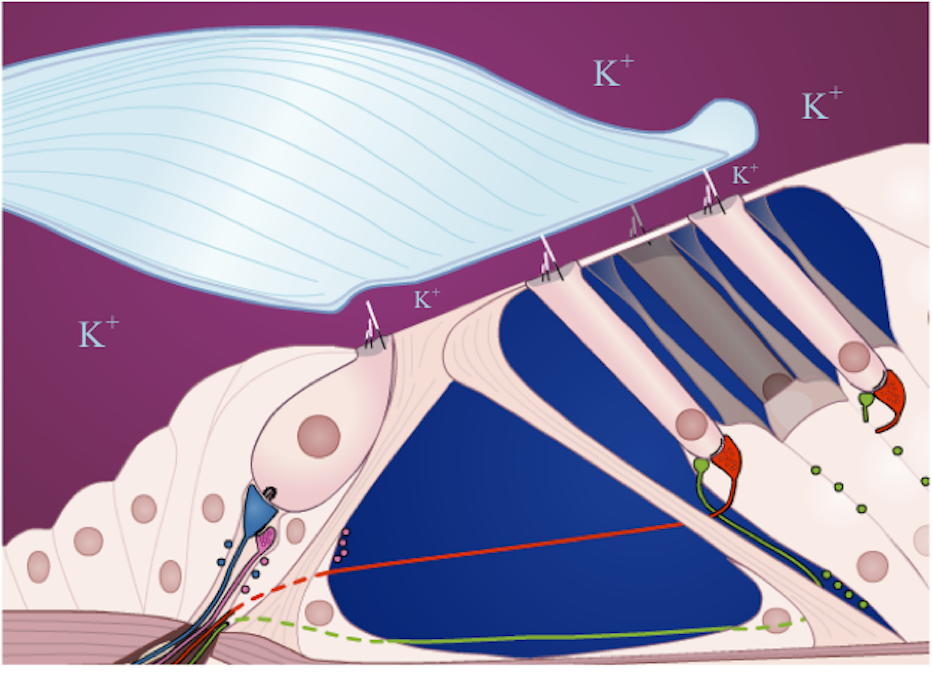
[Fig. 2.12] Détail de l'organe de Corti
(Cliquez pour voir l'original)
On distingue deux types de cellules ciliées : les cellules ciliées internes et les cellules ciliées externes. Leur anatomie est très semblable. Toutefois leur rôle est différent, tout comme leur disposition dans l’organe de Corti.
a. Les cellules ciliées internes : anatomie et généralités
Les cellules ciliées internes (CCI) sont les cellules situées vers l’extrémité interne de l’organe de Corti. En tant que cellules sensorielles, leur extrémité inférieure est directement reliée à deux neurones par des synapses, qui permettent la transduction de l’onde en signal électrique. Elles sont rangées en une seule ligne tout le long de la cochlée – soit près de 3500 au total, et sont en moyenne les plus sollicitées lors du processus de l’audition. En effet, pas moins de dix liaisons synaptiques sont présentes en moyenne par CCI, ce qui représente environ 95% du nerf auditif cochléaire, responsable de la transmission du signal électrique au cerveau.
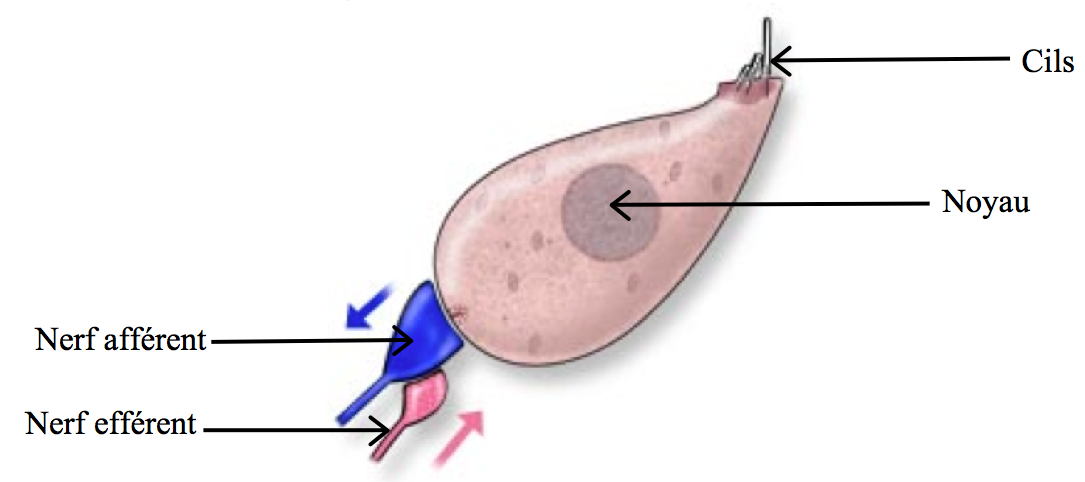
[Fig. 2.13] Schéma d'une cellule ciliée interne
(Cliquez pour voir l'original)
Au sommet de la CCI se trouvent des cils (les stéréocils) alignés par ordre croissant de taille : le plus petit vers l’intérieur de l’organe de Corti, et le plus grand vers l’extérieur. Il est à noter que les stéréocils ne se régénèrent pas après une détérioration, ce qui se traduit généralement par une surdité partielle dans une gamme de fréquence donnée.
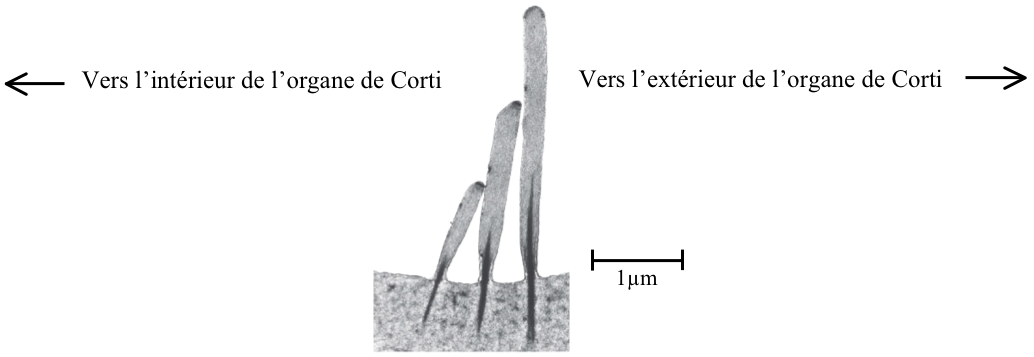
[Fig. 2.14] Vue des cils d'une cellule ciliée interne au MET
(Cliquez pour voir l'original)
Sur chacun d’entre eux se trouve un canal ionique, de la forme d’une porte, fermée en temps normal. Celui-ci relie chaque stéréocil à son plus grand voisin par un filin moléculaire appelé lien apical (voir figure 2.15).
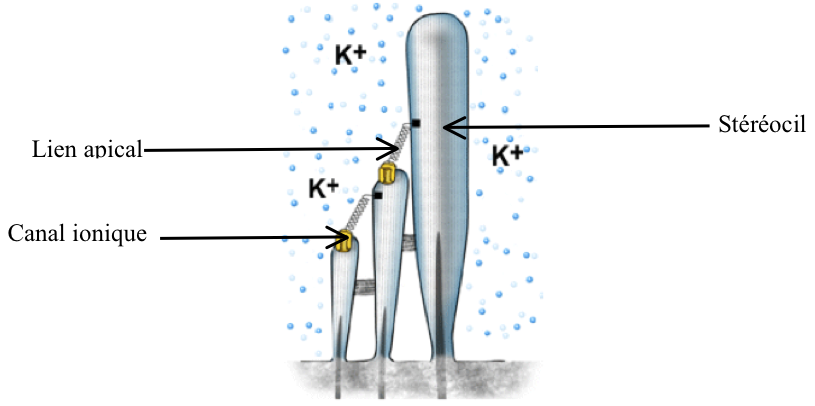
[Fig. 2.15] Détail de l'anatomie des stéréocils
(Cliquez pour voir l'original)
b. Les cellules ciliées internes : rôle et fonctionnement
Lors du passage d’une onde dans la cochlée, la membrane basilaire oscille de haut en bas, et stimule l’organe de Corti : les cils des CCI s’inclinent. Leur inclinaison provoque la tension des liens apicaux qui ouvrent les canaux ioniques. Ceux-ci étant ouverts, les ions K+ pénètrent dans la CCI et la parcourent jusqu’à son extrémité inférieure, où ils provoquent l’activation de la synapse du nerf afférent. Celle-ci induit une impulsion bioélectrique dans le nerf, qui la transmet jusqu’au cerveau (voir figure 2.16).
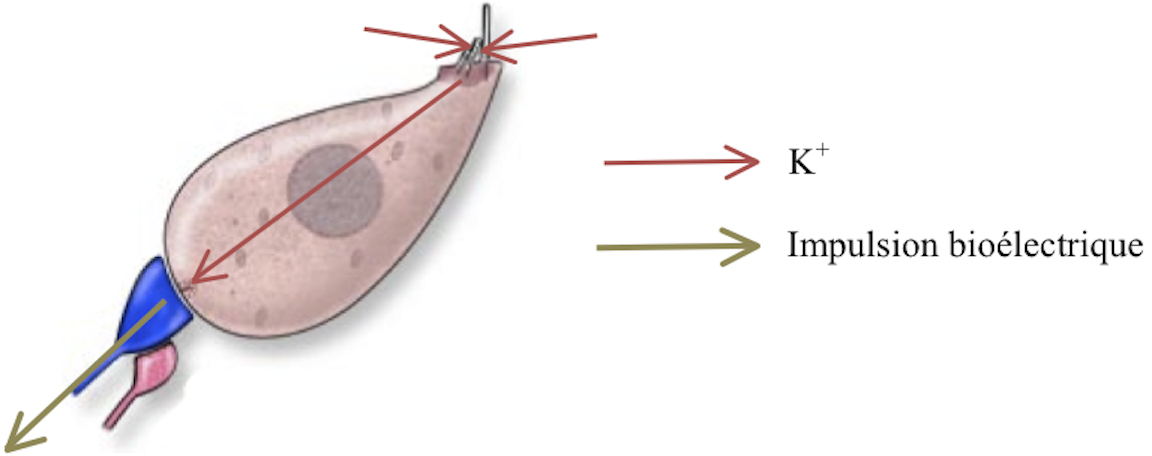
[Fig. 2.16] Schéma de la transduction au niveau d'une CCI
(Cliquez pour voir l'original)
c. Les cellules ciliées externes : anatomie et généralités
Les cellules ciliées externes (CCE) sont des cellules situées vers l’extérieur de l’organe de Corti. Au même titre que les cellules ciliées internes, ce sont des cellules sensorielles. Elles sont donc également reliées à des nerfs au niveau de leur extrémité inférieure.
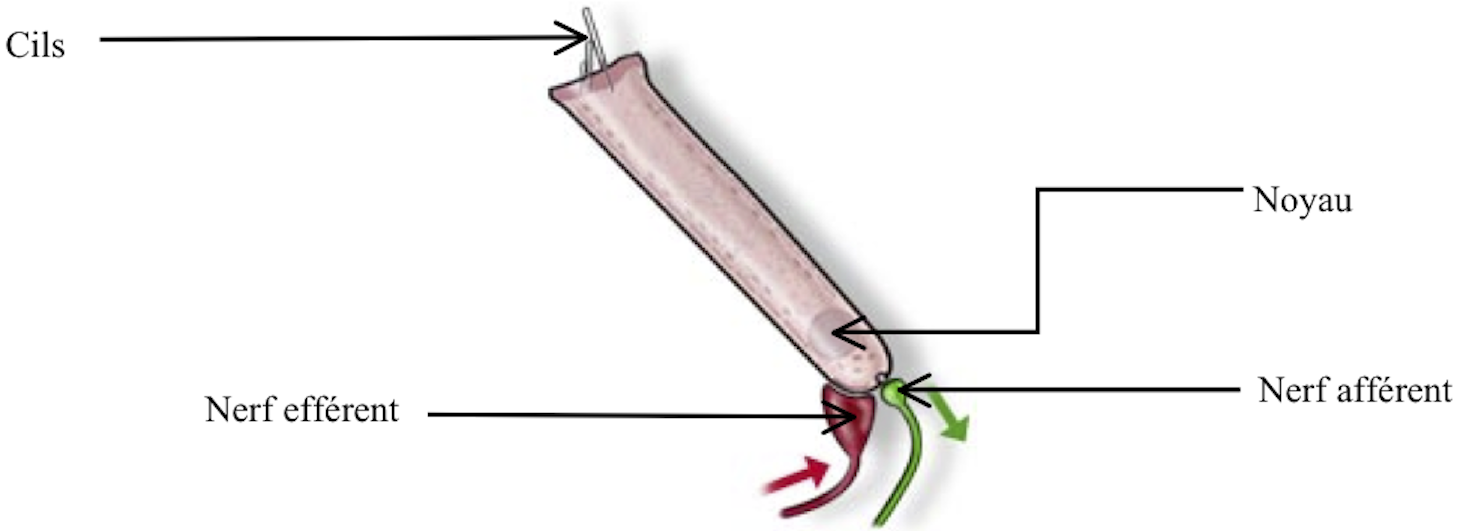
[Fig. 2.17] Schéma d'une cellule ciliée externe
(Cliquez pour voir l'original)
En revanche, leur disposition dans l’organe de Corti est quelque peu différente de celle des CCI. En effet, elles sont rangées en 3, 4 ou 5 rangées selon la zone de la cochlée où elles se trouvent (voir figure 2.18). Nous verrons dans une prochaine partie que cette répartition respecte la tonotopie cochléaire.
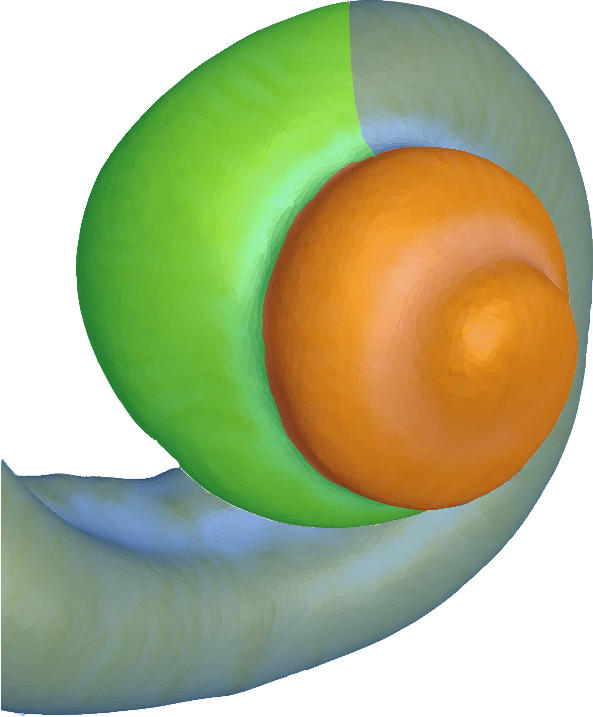
[Fig. 2.18] Schéma des zones de répartition des CCE dans la cochlée
(Cliquez pour voir l'original)
Légende
- : 3 rangées
- : 4 rangées
- : 5 rangées
Ce sont donc au total près de 12000 CCE qui se répartissent le long de l’organe de Corti, soit près de 4 fois plus que le CCI. Pourtant, malgré ce nombre important, les terminaisons nerveuses qui en partent ne représentent que 5% du nerf cochléaire. Ce contraste s’explique par le rôle des CCE, différent de celui des CCI.
d. Les cellules ciliées externes : rôle et fonctionnement
Afin de comprendre le rôle des CCE, il faut préciser le fonctionnement des CCI. En effet, les CCI ne sont pas en contact direct avec la membrane tectoriale, et il faut donc que la membrane basilaire vibre suffisamment pour les mettre en contact et activer le processus de transduction. Cette vibration correspond au passage d’une onde d’au moins 50 dB. En dessous de ce seuil, les CCI ne sont normalement pas stimulées.
C’est là que rentrent en jeu les CCE. Ce sont des amplificateurs naturels des vibrations de la membrane basilaire, pour des ondes de moins de 50 dB. Leur fonctionnement est globalement le même que celui des CCI : lorsqu’une onde sonore passe dans la cochlée, les cils des CCE s’inclinent et leur canaux ioniques s’ouvrent.
La différence réside dans le traitement du signal par le cerveau, envoyé après la transduction. En effet, celui-ci va provoquer un réflexe de rétrocontrôle : un nouveau signal électrique sera envoyé aux CCE via leurs nerfs efférents, ce qui provoquera leur contraction. Ce processus est appelé électromotilité.
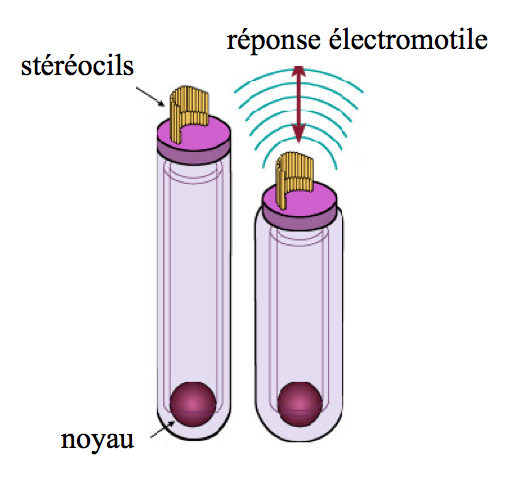
[Fig. 2.19] Électromotilité d'une cellule ciliée externe
(Cliquez pour voir l'original)
La contraction des CCE accentue la vibration de la membrane basilaire, qui rentre alors en contact avec les CCI, où la transduction s’effectue.
5) Décomposition d'une onde
a. Tonotopie cochléaire
La tonotopie, du grec tonos, le ton et topos, le lieu, est la répartition de la sensibilité d’un milieu à différents sons. Appliqué à la cochlée, cela révèle l’amplitude de la vibration de la membrane basilaire en fonction de la fréquence d’une onde. C’est en déroulant la cochlée (figure 2.20) qu’on arrive à définir sa tonotopie (figure 2.21).

[Fig. 2.20] Schéma du déroulage d'une cochlée
(Cliquez pour voir l'original)
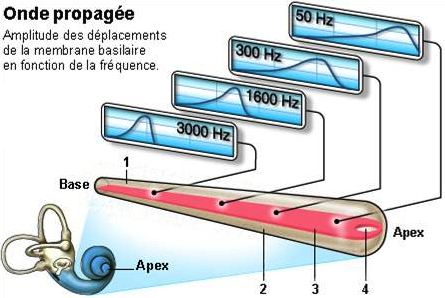
[Fig. 2.21] Représentation de la tonotopie cochléaire passive
(Cliquez pour voir l'original)
On remarque ainsi que les ondes dont les fréquences sont les plus élevées (sons aigus) sont principalement captées par la base de la cochlée, tandis que celles dont la fréquence est plus basse (sons graves) sont plutôt captées par l’extrémité de la cochlée (vers l’apex).
Cette tonotopie qui n’exige pas de contrôle moteur est appelée tonotopie passive, ou mécanisme passif.
Il existe également une tonotopie dite tonotopie active, ou mécanisme actif, qui dépend quant à elle de l’action des cellules ciliées externes. Pour un son pur de 10 kHz, par exemple, la tonotopie active induit une amplification d’environ 50 dB sur une portion très fine de la membrane basilaire (voir figure 2.22). Il va de soi que cette portion va varier en fonction de la fréquence du son reçu.
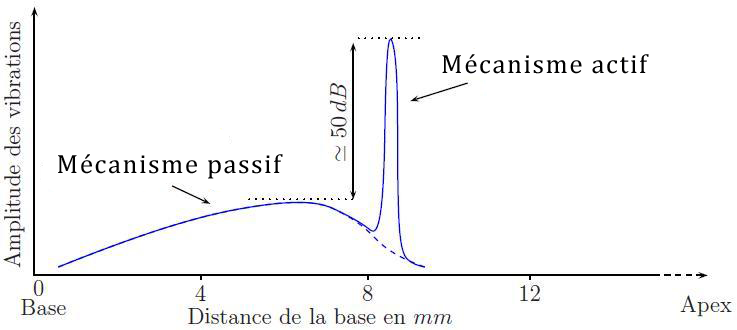
[Fig. 2.22] Représentation de la tonotopie cochléaire pour un son pur de 10kHz
(Cliquez pour voir l'original)
b. Décomposition d'une onde
Les sons entendus quotidiennement sont généralement complexes. Ils sont donc formés de plusieurs ondes additionnées entre elles (voir la partie I). Pourtant, un son complexe n’est reconstitué, et donc analysé comme tel, qu’au niveau du cerveau. En effet, l’oreille interne va le décomposer et envoyer au cerveau un signal bioélectrique correspondant à chacune de ses composantes.
Pour aborder le codage électrique d’un son complexe, il faut donc considérer le codage de chacune des composantes de l’onde sonore associée, ce qui revient à étudier le codage d’un son pur (voir figure 2.23).
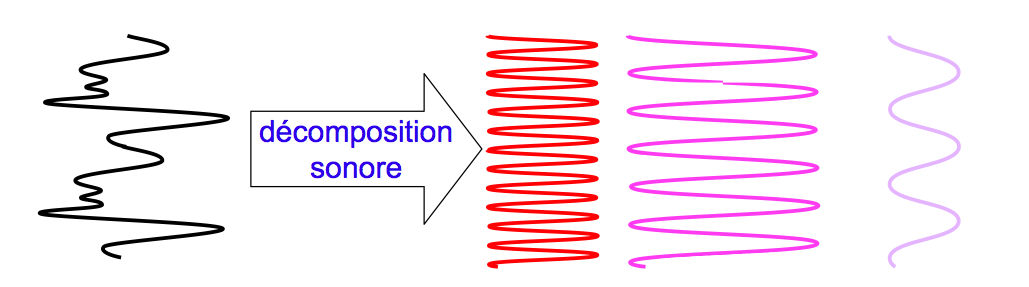
[Fig. 2.23] Schématisation d'une décomposition sonore
(Cliquez pour voir l'original)
Un tel codage se fait au travers de deux paramètres : la fréquence de l’onde et son intensité, ou amplitude.
La fréquence est codée en fonction des cellules ciliées internes qui envoient un signal, suivant la tonotopie cochléaire. Un signal des CCI de la base de la cochlée codera donc pour une haute fréquence, et inversement pour un signal des CCI proches de l’apex (voir figure 2.24).

[Fig. 2.24] Représentation des fréquences codées par les CCI en fonction de la situation dans la cochlée
(Cliquez pour voir l'original)
L’intensité est codée en fonction de la fréquence de décharge du signal émis par les CCI stimulées. La fréquence de décharge correspond au nombre d’impulsions électriques envoyées par les CCI en une seconde. C’est un taux proportionnel à l’inclinaison des CCI : plus elles s’inclinent, plus la transduction est mobilisée, et plus le nombre d’impulsions générées est important (voir figure 2.25).
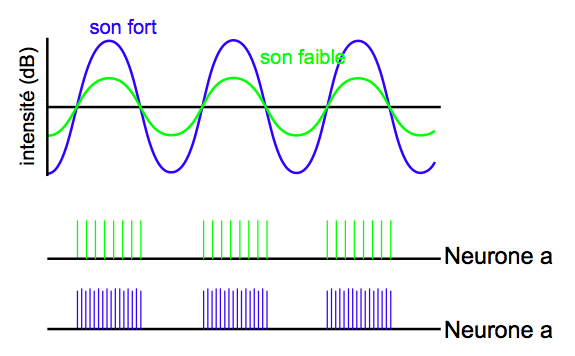
[Fig. 2.25] Fréquence de décharge d'un même neurone pour deux sons d'intensités différentes
(Cliquez pour voir l'original)
À l’aide de ces deux mécanismes de codage, l’oreille interne décompose un son et le transmet au cerveau par l’intermédiaire du nerf cochléaire.

